
摘要
动脉粥样硬化性心血管疾病导致心脏病发作和中风,这是世界范围内死亡的主要原因1。当低密度脂蛋白与硫酸肝素蛋白聚糖(HSPGs)结合2并被困在大、中动脉的内皮下空间时,动脉粥样硬化斑块的形成就开始了,从而导致动脉壁的慢性炎症和重塑2。增殖诱导配体是一种与HSPGs3结合的细胞因子,但这种相互作用的生理机制在很大程度上是未知的。本研究表明,基因消融或抗体介导的APRIL耗竭会加重小鼠动脉粥样硬化。在机制上,我们证明了APRIL通过与硫酸肝素蛋白聚糖2 (HSPG2)的硫酸肝素链结合来保护动脉粥样硬化,这限制了低密度脂蛋白的保留,巨噬细胞的积累和坏死核心的形成。事实上,在表达硫酸肝素缺乏的HSPG2的小鼠中,抗体介导的四月耗竭对动脉粥样硬化的发展没有影响。用一种特异性抗APRIL抗体治疗可促进APRIL与HSPGs结合,从而减少实验性动脉粥样硬化。此外,与HSPGs结合的一种人类APRIL蛋白的血清水平,我们称之为非典型APRIL (nc-APRIL),与动脉粥样硬化患者的长期心血管死亡率独立于传统危险因素。我们的数据揭示了APRIL的特性对血管稳态具有广泛的病理生理意义。
这是订阅内容的预览,可通过您的机构访问
访问选项
访问Nature和其他54种Nature组合期刊
获取自然+,我们最具价值的在线订阅
99€/ 30天
随时取消
中国客户的订阅信息
我们有一个专门的网站为我们的中国客户。请登录naturechina.com订阅本刊。
购买这篇文章
- 在Springer Link上购买
- 即时访问全文PDF
价格可能会在结账时计算当地税收




类似的内容被其他人浏览

数据可用性
RNA测序数据集(来自血管平滑肌细胞)可在Gene Expression Omnibus中获得,加入代码为GSE117963和GSE17858。如有合理要求,可向通讯作者索取所有其他相关数据。本文提供了源数据。
代码的可用性
临床研究使用SPSS(20.0版本,SPSS Inc.)进行计算,R版本为3.6.0 (https://www.R-project.org/),使用R软件包survival (https://CRAN.R-project.org/package=survival)和survminer (https://CRAN.R-project.org/package=survminer)进行生存分析。对于质谱数据的分析,获取的原始数据文件使用Proteome Discoverer 2.4.1.15 SP1处理DDA实验数据,或使用Skyline 20.1.0.155处理PRM实验数据,或使用Mascot 2.3.02 (Matrix Science, London, UK)和Phenyx (GeneBio, Geneva, Switzerland)作为搜索引擎。RNA-seq数据使用FastQC v0.11.3 (https://www.bioinformatics.babraham.ac.uk/projects/fastqc/)进行质量控制,并使用Trim Galore v0.4.1包装器(https://www.bioinformatics.babraham.ac.uk/projects/trim_galore/)进行裁剪。使用Tophat v2.0.12将Reads与GRCm38小鼠参考基因组进行比对。将最低地图质量为20的Reads导入Seqmonk 1.45.4 (http://www.bioinformatics.babraham.ac.uk/projects/seqmonk)。
参考文献
动脉粥样硬化的变化。自然,592,524-533(2021)。
陈志强,陈志强。动脉粥样硬化的免疫学研究。[j] .中国生物医学工程学报,2016,33(5):387 - 387(2017)。
英戈尔德,K.等。蛋白聚糖作为april特异性结合伙伴的鉴定。中华医学杂志,2011,33(5):387 - 398(2005)。
Vincent, F. B, Morand, E. F, Schneider, P. & Mackay, F. BAFF/APRIL系统在SLE发病机制中的作用。中华风湿病杂志,2014,31(5):357 - 357。
Castigli, E.等。april缺陷小鼠中IgA类转换受损。自然科学进展。美国101,3903-3908(2004)。
等。由中性粒细胞分泌的APRIL与硫酸肝素蛋白聚糖结合,在人粘膜中形成浆细胞壁龛。j .中国。《投资管理》,2008,28(2)。
mcmccarron, M. J, Park, P. W. & Fooksman, D. R.。CD138通过调节成熟浆细胞的存活来调节其选择。血液129,2749-2759(2017)。
Hymowitz, s.g.等。april受体复合物的结构:与BCMA一样,TACI仅利用单一的富含半胱氨酸的结构域进行高亲和力配体结合。生物。化学280,7218-7227(2005)。
桑德伯格等。肿瘤坏死因子超家族配体APRIL (TNFSF13)在血小板活化后释放并在动脉粥样硬化中表达。Thromb。中国医学杂志,2009,33(4):391 - 391。
主啊,m.s.等。perlecan在纤维化中的多重作用。生物工程学报,36(2):444 - 444(2018)。
trans - lundmark, K.等。肝磷脂中的硫酸肝素促进小鼠动脉粥样硬化:在脂质通透性、脂质保留和平滑肌细胞增殖中的作用。Circ. Res. 103,43 - 52(2008)。
Sarrazin, S, Lamanna, W. C. & Esko, J. D.硫酸乙酰肝素蛋白聚糖。冷泉港。教谕。中国生物医学工程学报,2011,49(2):481 - 481。
硫酸肝素在炎症中的作用。免疫学杂志,6,633-643(2006)。
Bernelot Moens, S. J.等。B细胞生长因子APRIL对动脉粥样硬化斑块定性和免疫学特性的影响。《科学通报》,2016,33(5):481 - 481。
Haselmayer, P., Vigolo, M., Nys, J., Schneider, P. & Hess, H.系统性红斑狼疮小鼠模型对可溶性TACI的反应优于可溶性BAFFR,这与浆细胞的消耗有关。欧元。[j] .中华免疫学杂志,2017,37(5):559 - 567。
Tsiantoulas等人。B细胞活化因子中和会加重动脉粥样硬化。流通138,2263-2273(2018)。
Tsiantoulas等人。血浆IgE升高加速分泌型IgM缺乏的动脉粥样硬化。Circ. Res. 120,78 - 84(2017)。
迪什曼,a.f.等。一种变质蛋白中折叠开关的进化。科学,371,86-90(2021)。
Schillinger, M.等。炎症和颈动脉粥样硬化风险研究(ICARAS)。循环111,2203-2209(2005)。
温克尔曼,b.r.等。LURIC研究的基本原理和设计-功能基因组学、药物基因组学和心血管疾病长期预后的资源。药物基因组学2(增刊1),S1-S73(2001)。
Puymirat, E.等。急性心肌梗死:1995年至2015年FAST-MI项目(法国急性st段抬高或非st段抬高心肌梗死登记处)20年间患者特征、管理和6个月结局的变化。发行量136,1908-1919(2017)。
Kowalczyk-Quintas, C.等。诱导外胚层发育不良的功能阻断抗外胚层发育不良蛋白A (EDA)单克隆抗体的产生和鉴定。生物。化学,289,4273-4285(2014)。
周明英等。氧化特异性表位是小鼠和人类先天天然抗体的主要靶点。j .中国。科学通报,2009,(5):444 - 444。
金佳,S., Vázquez, A. M., Levin, M., borsamen, J., Fogelstrand, P.血管干预引起的内膜增生导致脂蛋白滞留和动脉粥样硬化加速。杂志。众议员5,e13334(2017)。
Tom, R., Bisson, L.和Durocher, Y.。用线性PEI转染HEK293-EBNA1细胞制备重组蛋白。冷泉港。Protoc。https://doi.org/10.1101/pdb.prot4977(2008)。
Schindelin, J.等。斐济:生物图像分析的开源平台。Nat. Methods 9,676 - 682(2012)。
博森,C.等。与BAFF- r不同,TACI仅被低聚BAFF和APRIL激活,以支持活化的B细胞和质母细胞的存活。《中国医学杂志》(英文版),2008。
Kowalczyk-Quintas, C.等。抗BAFF抗体Belimumab对膜结合BAFF的抑制作用前面。免疫学杂志,9,26(2018)。
Schneider, P., Willen, L. & Smulski, C. R.研究配体-受体相互作用和TNF超家族成员受体激活的工具和技术。方法:中国生物医学工程学报(英文版)。
Manza, l.l., Stamer, s.l., Ham, a.j.。L., Codreanu, s.g. & Liebler, d.c.样品制备和消化蛋白质组学分析使用自旋过滤器。蛋白质组学5,1742-1745(2005)。
Wiśniewski, J. R., Zougman, A., Nagaraj, N. & Mann, M.蛋白质组分析的通用样品制备方法。Nat. Methods 6,359 - 362(2009)。
Rappsilber, J, Ishihama, Y. & Mann, M.停止和走提取技巧的基质辅助激光解吸/电离,纳米电喷雾,和蛋白质组学的LC/MS样品预处理。分析的。化学75,663-670(2003)。
奥尔森,j.v.等。通过锁定质量注入到c -捕集器的Orbitrap质谱仪的百万分之一质量精度。摩尔。细胞。蛋白质组学,2010-2021(2005)。
Hellings, W. E., Moll, F. L, de Kleijn, D. P.和Pasterkamp, G. 10年的动脉粥样硬化快速研究经验。Cardiovasc。成岩作用。[j] .农业学报,2012,63-73。
梅耶,F. J.等人。炎症状态和颈动脉粥样硬化的联合影响:一项12年随访研究。Stroke 47,2952 - 2958(2016)。
Battle, A., Brown, C. D., Engelhardt, B. E.和Montgomery, S. B.基因对人体组织基因表达的影响。自然,550,204-213(2017)。
致谢
我们感谢A. Fabry和T. Wenko在体内实验研究方面的帮助,以及C. Friedl在共聚焦显微镜方面的帮助。这项工作得到了奥地利科学基金(SFB F54)、欧盟(FP7 VIA)和Leducq基金会(TNE-20CVD03)对C.J.B的资助,以及欧洲心血管疾病研究区网络(I4647)和英国心脏基金会(RCAG/917)对d.t.d.t.的资助,奥地利科学基金(I4963)也支持这项工作。瑞士国家科学基金资助项目(31003A_176256, 310030E_197000)。z.m由英国心脏基金会(RCAM/104, RCAM-659, RRCAM.163),英国心脏基金会卓越研究中心(RE/18/1/34212), NIHR剑桥生物医学研究中心(RG85315),欧盟(FP7 VIA;RCAG/430)和欧洲研究理事会(ERC)。T.H.由NWO Veni资助(91619012)。
作者信息
作者及单位
贡献
D.T.构思和设计了这项研究,进行了大部分实验,分析和解释数据,并撰写了手稿。M.E.和P.S.生成了材料,进行了表征nc-APRIL的实验并解释了数据。于G.O。,S.K L.W,实际高度,T.H。M.G.K M.O.-K。, l.g., f.p., J.E.M.和P.F.协助小鼠研究并为实验提供技术支持。M.C.辅助免疫荧光分析。D.S.协助小鼠研究。J.L.和H.F.J.提供了小鼠VSMCs的RNA-seq数据。A.M.和J.W.进行了质谱分析并分析了数据。F.J.M.和F.F.分别对ICARAS和LURIC临床数据进行统计分析。O.D.和J.B.为实验提供了试剂和技术专长,并对手稿进行了严格的修改。M.H.参与了人体样本的分析。T.S和N.D.提供FAST-MI临床研究的样本,并对数据进行统计分析。h.s., W.M.和Z.M.提供了来自LURIC临床研究的样本,并测量了nc-APRIL水平。gp提供了材料。H.H.提供了材料,并对手稿进行了严格的修改。z.m和P.S.参与了研究设计,解释了数据并批判性地修改了手稿。C.J.B.设计了这项研究,解释了数据并撰写了手稿。
相应的作者
道德声明
相互竞争的利益
d.t., c.j.b., P.S.和M.E.在一项专利申请(EP20217536.0;维也纳医科大学(奥地利)和奥地利科学院分子医学研究中心(奥地利)提交了一份申请,将c-APRIL和nc-APRIL用于心血管疾病的诊断和治疗。O.D.是Adipogen生命科学公司的员工,为本研究提供了部分试剂。
额外的信息
《自然》杂志感谢Peter Libby、William Sessa和其他匿名审稿人对本文同行评议的贡献。
施普林格·自然对已出版地图的管辖权要求和机构关系保持中立。
扩展数据图和表
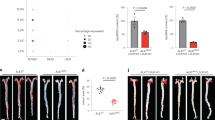
图1 APRIL缺乏不改变斑块平滑肌细胞和胶原蛋白含量,或循环单核细胞、B或T淋巴细胞的数量Ldlr−−/老鼠。
Ldlr−/−Tnfsf13+/+或Ldlr−/−Tnfsf13−/−小鼠喂食致动脉粥样硬化饮食10周。a,全体重、血浆甘油三酯水平和面部病变大小(n = 10 Ldlr - / - Tnfsf13+/+小鼠,n = 12 Ldlr - / - Tnfsf13 - / -小鼠)。b - d,主动脉起源(左)的DAPI- (b)、α-SMA- (c)和天狼星红染色病变(d)的代表性显微照片,以及显示平均脱细胞(b;n = 9只Ldlr - / - Tnfsf13+/+小鼠,n = 12只Ldlr - / - Tnfsf13 - / -小鼠,P = 0.036), α- sma阳性(c, n = 9只Ldlr - / - Tnfsf13+/+小鼠,n = 10只Ldlr - / - Tnfsf13 - / -小鼠),胶原阳性面积归一化(d;n = 10只Ldlr−/−Tnfsf13+/+小鼠,n = 12只Ldlr−/−Tnfsf13−/−小鼠)。e - g,脾CD3+、CD4+(定义为CD3+CD4+CD8−)和CD8+ T细胞(定义为CD3+CD8+CD4−)绝对数量(e)、腹膜B-1a(定义为B220lowCD11bintCD5−)、B-1b(定义为B220lowCD11bintCD5−)和CD23+ B-2(定义为B220highCD11b−CD5−CD23+)细胞频率(f)和外周血循环ly6high、Ly6Cint和Ly6Clow单核细胞频率(g)的代表性流式细胞术点图(上)和定量图(下)。所有结果均为均值(双尾非配对学生t检验)。标尺,200 μm。
扩展数据图2 BCMA对于动脉粥样硬化的发展是不可或缺的。
经致死性照射的Ldlr - / -小鼠注射来自Bcma+/+ (hem-Bcma+/+)或Bcma - / -供体(hem-Bcma - / -)的骨髓,喂食致动脉粥样硬化饮食10周。a,脾脏中的Bcma mRNA (n = 11 hem-Bcma+/+, n = 13 hem-Bcma - / -小鼠)。b, h&e染色主动脉根部病变的代表性显微照片(左)和主动脉根部病变的平均大小(右),以μm2 /切片表示(n = 11 hem-Bcma+/+, n = 12 hem-Bcma−/−小鼠)。c,全身体重、血浆甘油三酯水平和面部病变大小(n = 11 hem-Bcma+/+, n = 13 hem-Bcma−/−小鼠)。d,血浆总胆固醇(n = 11 hem-Bcma+/+, n = 13 hem-Bcma−/−小鼠)。e、FO/T2、MZ、CD21+CD23−(P = 0.028)、T1 (P = 0.023)、NF (P = 0.031)和B-1 B细胞的绝对数量(n = 11只hem-Bcma+/+, n = 13只hem-Bcma−/−)。f, CD3+、CD4+和CD8+ T细胞的绝对数目(n = 11只hem-Bcma+/+, n = 13只hem-Bcma−/−小鼠)。g, h,腹膜B-1a (P = 0.003)、B-1b (P = 0.021)和CD23+ B-2细胞的频率(P = 0.0003) (g)和总IgM (P = 0.002)、IgG1、IgG2b (P = 0.002)、IgG2c (P = 0.001)、IgG3 (P = 0.032)和IgA血浆抗体滴度(h);n = 11 hem-Bcma+/+, n = 13 hem-Bcma - / -小鼠)。结果均为mean±s.e.m, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001(双尾Mann-Whitney u检验或双尾unpaired Student 's t检验)。标尺,200 μm。
扩展数据图3四月是由小鼠和人类VSMCs产生的。
a、基因型-组织表达(GTEx)项目中TNFSF13基因在人体组织中的表达GTEx项目得到了美国国立卫生研究院主任办公室共同基金以及NCI、NHGRI、NHLBI、NIDA、NIMH和NINDS的支持。本文中描述的数据于2021年1月21日从GTEx门户网站获得,dbGaP登录号为phs000424.v8.p2。结果显示中位数(主动脉:中位数= 55.05,n = 432;冠状动脉:中位数= 40.7,n = 240)。b, c,主动脉弓(AA)和胸降主动脉(DT) VSMCs的Bulk RNA-seq分析(b;n = 3-5只小鼠)(GSE117963)和分离后保存在Trizol中或培养4-5代直至分析的小鼠原代VSMCs (c;GSE17858)。表达TNFSF13、MYH11和KI67基因。d、重组人TNF、天然人LDL或人oxLDL四次重复刺激人脐动脉平滑肌细胞的TNFSF13和IL6基因表达(TNF刺激是三个独立实验的代表;P = 0.003)。结果均值±s.e.m. **P < 0.01(单因素方差分析和Tukey检验)。
扩展数据图4四月结合HSPG2。
a,在肝素存在或不存在的情况下,用人或小鼠Flag-APRIL孵育的HUVECs的代表性显微照片(左)和定量(右),用FITC偶联的抗flag M2抗体进行染色,并通过共聚焦显微镜进行分析。b,用Flag-APRIL或flag标记的细菌碱性磷酸酶(Flag-BAP)孵育并用抗flag M2抗体染色的HUVECs的流式细胞术分析。c,通过琼脂糖珠偶联抗flag M2抗体进行下拉实验,鉴定HUVEC培养中APRIL的蛋白结合伙伴,然后进行质谱分析。d,通过ELISA定量测定小鼠基底膜中APRIL与被包被HSPGs的结合(****P < 0.0001,双尾未配对学生t检验)。所示数据至少代表了两个独立实验(a, b, d)。e, f,小鼠颈动脉切片的显微照片,用小鼠多聚体Flag-APRIL孵育,用与PE偶联的抗flag抗体或与PE偶联的抗小鼠APRIL生物素化抗体(2C8)和链亲和素(e;刻度条,75 μm,数据来源于两个独立的实验),或用小鼠多聚体flag -四月在肝素存在或不存在的情况下,用抗四月生物素化抗体(2C8)或抗hspg2或仅二抗(仅2)染色(f;比例尺,75 μm,数据来源于一次实验)。g、定量表面等离子体共振(Biacore)分析可溶性人Fc-APRIL (total)、人典型Fc-APRIL (human Fc-c-APRIL)、人非典型Fc-APRIL (human Fc-nc-APRIL)、小鼠典型Fc-APRIL (mouse Fc-c-APRIL)、小鼠非典型Fc-APRIL (mouse Fc-nc-APRIL)以及阴性对照EDAR-Fc和人Fas-Fc对链亲和素传感器芯片A偶联的生物素化肝素的亲和力(n = 3个独立实验)。所有结果均为平均值±s.e.m。IntDen,综合密度。
图5抗APRIL抗体108、2C8和Apry-1-1对小鼠APRIL具有特异性。
a,还原条件下抗mapril mAb 108和2C8的考马斯蓝分析。b、抗mapril 108、Apry-1-1和2C8的Fc部分的同型分析。纯化后的抗体包被在ELISA板上,显示有针对不同同型的过氧化物酶结合抗体。c,与TACI-Fc相比,108和Apry-1-1对人和小鼠apry -1的抑制活性。在固定、不饱和浓度为108、Apry-1-1或TACI-Fc的情况下,在BCMA-Fas报告细胞上滴定Flag-human APRIL和Flag-mouse APRIL的两个剪接变体(±Ala112)。数据显示,108和Apry-1-1对mAPRIL的两个剪接变异体都有大致的抑制作用,但与人类APRIL没有交叉反应。d, 2C8与小鼠APRIL结合的ELISA检测。用过氧化物酶偶联抗小鼠抗体评价2C8与人fc -小鼠APRIL包被板的结合。a, b,数据是两个独立实验的代表。
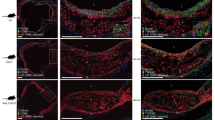
图6 APRIL竞争LDL与蛋白聚糖的结合。
a,在小鼠多聚体Flag-APRIL存在或不存在的情况下,用共聚焦和荧光显微镜分析抗载脂蛋白ob抗体结合小鼠颈动脉切片的代表性显微照片(左)和定量(右)(不含APRIL, n = 7;四月,n = 9;P = 0.0004)。b,结合人LDL的量(三次);在不同量的人重组APRIL存在下,HEK293野生型细胞表面的表达量(以流式细胞术定量)。c,在喂食致动脉粥样硬化饮食10周的Ldlr - / - Tnfsf13+/+或Ldlr - / - Tnfsf13 - / -小鼠(n = 8 Ldlr - / - Tnfsf13+/+小鼠,n = 12 Ldlr - / - Tnfsf13 - / -小鼠)的主动脉起源apob染色病变(左)和apob阳性归一化为DAPI+病变区域(右)的代表性显微照片;P = 0.035)。所示数据来自4个独立实验,每组7 - 9个部分(a),代表3个独立实验(b)。所有结果均为mean±s.e.m., *P < 0.05, ***P < 0.001(双尾未配对学生t检验)。比例尺,20 μm (a)和200 μm (c)。
图7阻断性抗四月抗体Apry1-1治疗不改变B细胞、IgM和血浆脂质水平Apoe−−/老鼠。
Apoe−/−小鼠每两周给予抗小鼠APRIL抗体(Apry-1-1)和对照ig(抗APRIL组),或TACI-Ig和同型IgG2b (TACI-Ig组),或同型IgG2b和对照ig(对照组)的混合物治疗,并在研究的最后8周喂食致动脉粥样硬化饮食。a - f,点阵图显示脾脏总B细胞(a)、卵泡(FO) B细胞(B)、边缘区(MZ) B细胞(c)的数量,以及腹膜B-1a (d)、腹膜B-1b (e)的频率和血浆中总IgM抗体水平(f)。g、野生型小鼠腹腔注射1 μg小鼠多聚体Flag-APRIL或1 μg Flag-APRIL与10 μg抗小鼠APRIL抗体(Apry-1-1)的混合物。注射后1、3、6 h采用ELISA法测定血浆中Flag-APRIL的含量(n = 4只小鼠Flag-APRIL, n = 5只小鼠Flag-APRIL + anti-APRIL (Apry1-1))。h,体重,血浆甘油三酯和胆固醇水平。a-f, h,所有结果均为平均值±s.e.m;(n = 10 Apoe−/−control, n = 12 Apoe−/−TACI-Ig, n = 10 Apoe−/−anti-APRIL)。***P < 0.001, ****P < 0.0001(单因素方差分析和Newman-Keuls检验)。
图8抗人APRIL抗体的表位定位,以及原生典型和非典型APRIL的大小不同,但由同一基因产生。
a-g,抗人APRIL抗体表位定位。a、Aprily1、2、3、5和10识别的表位通过截断的APRIL蛋白进行western blot定位。b-d、Aprily5和Aprily1或Aprily2识别不同的表位(d),而Aprily3和Aprily10识别与Aprily1、Aprily2和Aprily5不同的表位(e, f)。g。h, i,使用抗人APRIL抗体Aprily1, Aprily2, Aprily3, Aprily5, Aprily6, Aprily8, Aprily9, Aprily10, Mahya-1, 110.6,生物atacicept (TACI-Ig:受体TACI与Ig Fc区结合的重组融合蛋白,与APRIL结合)或阴性对照EctoD1,然后用c-APRIL特异性(h;ELISA 1)或nc- april特异性ELISA (i;ELISA 2)数据来源于此格式的一个实验。j-n,原生规范和非规范APRIL的大小不同。j, Flag-human APRIL(来自c-APRIL ELISA 1标准)在TACI-Fc(或TNFR2-Fc作为对照)和/或在Aprily2(或mIgG1作为对照)上被去除。然后用c-APRIL特异性(上)或nc-APRIL特异性(下)ELISA检测APRIL。k, 1, Flag-human APRIL(来自APRIL ELISA 1标准品)在固定化TACI-Fc或Aprily2上去除,然后通过大小排除色谱(SEC)对流动进行大小分离,并通过c-APRIL特异性(k;ELISA 1)或nc- april特异性(l;Elisa法;然后用酸洗脱TACI-Fc和Aprily2珠。m, n,对中和的洗脱液进行分级,用c-APRIL特异性(m)或nc-APRIL特异性(n) ELISA检测各组分中的APRIL。这些结果表明,Flag-c-APRIL具有3-mer的大小,而nc-APRIL要大得多。o, p,典型和非典型APRIL是由相同的TNFSF13基因位点产生的。利用CRISPR-Cas9技术灭活人巨噬细胞系U937中编码APRIL的TNFSF13基因。作为对照,TNFSF13B基因(编码BAFF)也被删除。用c-APRIL特异性(o)和nc-APRIL特异性(p) ELISA检测上清液中的APRIL。105、110、301和302描述了不同的克隆。
图9基于lc - ms的平行反应监测(PRM)分析纯化的人典型或非典型Fc-APRIL的胰蛋白酶消化。
a - C,使用Skyline软件分析原始数据,提取的产物离子色谱(XICs)以峰(上)或集成产物离子面积总和(下)的形式显示三种选定肽EEQYNSTYR (Fc部分)(a), LNLSPHGTFLGFVK(色氨酸C末端APRIL) (b)和LNLSPHGTFLGFVKL(错切色氨酸C末端APRIL) (C)。所选肽前体离子的MS2片段离子光谱示于右下角。尽管图a所示的肽是典型与非典型Fc-APRIL注射量比较的代表,但图c所示的c端错误切割的全色氨酸在非典型APRIL中是检测不到的。相对丰度以任意单位给出。右图为Fc-APRIL的FASTA序列,其中选定的色氨酸序列以蓝色或红色突出显示。注意b(109)和c(106)的不同音阶。d,人类C -APRIL的结构,强调了C末端对不同形式(规范和非规范)APRIL折叠的重要性。基于蛋白质数据库编号1XU1的表示突出了最后两个c端氨基酸(Lys232, Leu233)。还显示了TNF同源结构域(His98)和Asp142的n端氨基酸。所有这些残基在小鼠四月和人类四月中都是保守的,尽管在小鼠和人类四月中,围绕Asp142的序列是不同的。Leu233的c端羧基非常接近同一单体的His98(三个单体中分别为3.6 Å、4.1 Å和3.4 Å),也非常接近相邻单体的His98 (4.3 Å、3.8 Å和3.8 Å)。因此,His98和羧基Leu233似乎在四月的上表面形成了一个由六个盐桥组成的环。此外,Lys232与Asp142(在小鼠APRIL的3个单体中分别为4.3 Å、4.3 Å和5.7 Å)接触,与人APRIL的Asp142仅3.2 Å接触(PDB登录号4ZCH)。
权利和权限
关于本文

引用本文
Tsiantoulas博士,Eslami, M., Obermayer, G.等人。APRIL通过结合硫酸肝素蛋白聚糖限制动脉粥样硬化。自然,597,92-96(2021)。https://doi.org/10.1038/s41586-021-03818-3
收稿日期:2019年6月14日
录用日期:2021年7月12日
发布日期:2021年8月25日
发行日期:2021年9月2日
DOI: https://doi.org/10.1038/s41586 - 021 - 03818 - 3
这篇文章是由
-
动脉粥样硬化中细胞代谢失调:介质和治疗机会
Nature Metabolism (2024)
-
缺血血管损伤与修复中的铜沉降与缺铜
细胞凋亡(2024)
-
IFNγ与细胞外基质结合可防止致命的全身毒性
Nature Immunology (2023)
-
人硫酸肝素聚合酶复合物EXT1-EXT2的结构
自然通讯(2022)
-
动脉粥样硬化中针对炎症和免疫的治疗策略:如何进行?
Nature Reviews Cardiology (2022)
评论
通过提交评论,您同意遵守我们的条款和社区准则。如果你发现一些滥用或不符合我们的条款或指导方针,请标记为不适当。
